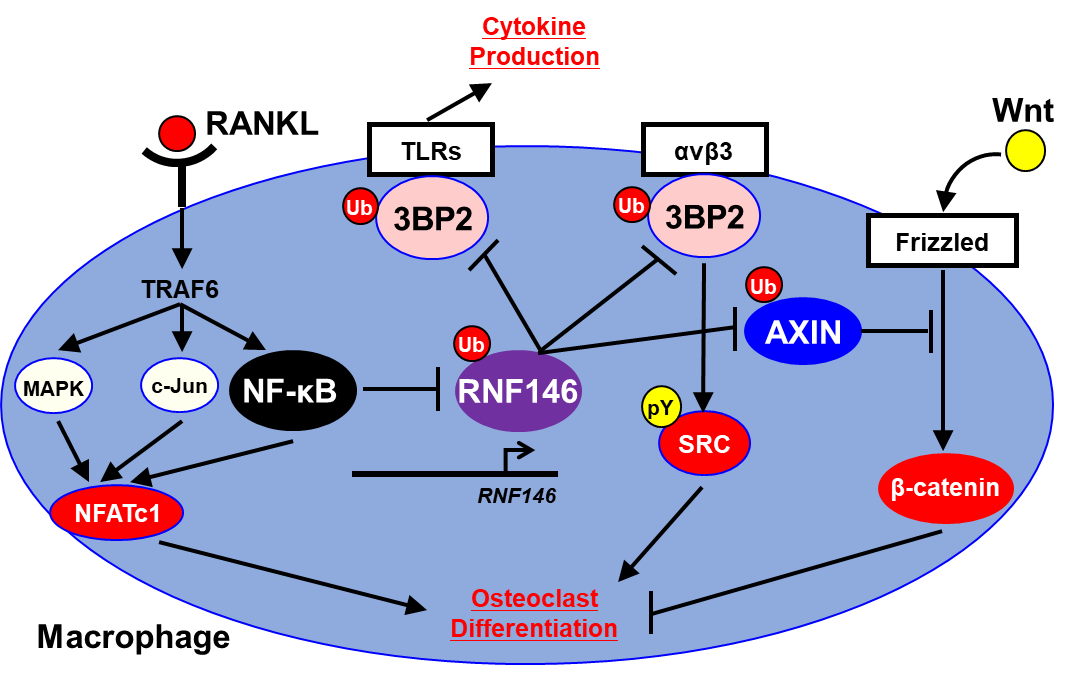
破骨細胞はサイトカイン“RANKL”の刺激により単球・マクロファージから分化して骨吸収を担い、骨粗鬆症や関節リウマチの骨破壊に関与することが知られている。RANKLによる破骨細胞必須転写因子NFATc1の発現増加は破骨細胞分化に必須のイベントと考えられている。確かにRANKL/NFATc1経路の活性化は破骨細胞分化に必須だが、一方で正常な破骨細胞の分化・機能にはインテグリン受容体の下流で起こるチロシンキナーゼSRC/Sykの活性化も重要である。逆に破骨細胞分化を負に制御するWnt/β-catenin経路は、RANKL刺激と同時に抑制されなければならない。一体その3つの経路をつなぐものは何か?私達は本論文で以下の如く、単球・マクロファージから破骨細胞への分化を制御する3つのシグナル経路を統合する因子がユビキチンリガーゼRNF146であることを明らかにした(図1)。
1. RANKLにより活性化される転写因子NF-κBがRNF146の転写を抑制する。
2. RNF146の発現低下により、その基質蛋白であるチロシンキナーゼ活性化因子“3BP2”やWnt制御因子“AXIN”の分解が抑制され、マクロファージ内でこれらの蛋白発現が増加する。
3. 3BP2の増加により、下流のSRC/Sykをはじめとするチロシンキナーゼが活性化され、破骨細胞分化・機能が促進する。
4. 一方AXIN増加に伴い、破骨細胞分化を負に制御するWnt/β-catenin経路は抑制され、破骨細胞分化・機能は更に促進する。
本論文では、破骨細胞分化の過程でRANKL誘導性NF-κBによるRNF146の抑制は必須であり、この現象により統合されたRANKL/NFATc1経路、インテグリン/3BP2/SRC/Syk経路、Wnt/β-catenin経路の3つが破骨細胞分化に向かって協調していくことを明らかにした。更に3BP2は、マクロファージにおいて発現量依存的にLPSへの感受性を増加させ、サイトカイン産生を亢進させることも明らかにし、RNF146は破骨細胞分化及び炎症の制御スイッチであることを示した。関節リウマチや炎症性腸疾患をはじめとする自己免疫性炎症疾患の病因を考える上で、3BP2制御に関わるRNF146の重要性が示唆された。
本研究は私の留学先であるUniversity of Toronto, Princess Margaret Cancer CenterのRobert Rottapel研究室で行いました。Prof. Rottapelは私と同じ膠原病内科医で、週1回Clinicで外来診療を、また月数回入院患者相談のオンコールもやっていましたが、多忙な診療の中で、基礎免疫学のみならず、骨代謝、癌、細胞骨格、バイオインフォマティクスなど全ての領域に精通し、世界に発信し続けているPhysician-scientistです。一生追いつけないであろう遙か遠い目標に出会えたことが、留学で得た最大の宝物でした。すぐそばにはT-cell receptorを発見したProf. Tak Makの巨大な研究室もあり、世界の研究のすごさや厳しさを学びました。それを岡山に戻って後進に伝えたいという思いで研究に取り組んでいます。結果の出なかった平凡な日本人研究者を5年半にわたり辛抱強く指導してくれたトロントの父、Robert Rottapel、そして長期に渡る私の留学を心より応援して下さった槇野博史岡山大学長、和田淳教授、大塚文男教授、佐田憲映教授をはじめとする岡山大学腎・免疫・内分泌代謝内科学の先生方に感謝致します。